
이후 서술되는 내용은 필자가 승현준(세바스찬 승) 교수님의 '커넥톰, 뇌의 지'를 읽고 인상 깊었던 부분을 정리한 것이다.
책에 소개된 내용에 추가적인 내용들을 덧붙여 작성하였다.
머리말
- 인간의 커넥톰은 경험에 따라 평생에 걸쳐 변화함 (4R)
- 재가중(Reweighting): 뉴런 간의 연결의 세기를 조정 (가중치를 변경)
- 재연결(Reconnection): 시냅스를 새로 만들거나 제거함
- 재배선(Rewiring): 가지가 자라거나 축소됨
- 재생성(Regeneration): 기존의 뉴런을 제거하고 완전히 새로운 뉴런을 생성
1부. 크기가 중요한가?
1장. 천재성과 광기
- 뇌의 크기와 지능 사이에는 상관관계는 있지만, 한 개인의 경우로 보면 관계가 없음 (그럴 가능성은 있겠지만)
- 피어슨 상관계수(Pearson’s correlation coefficient): 상관관계의 강도를 1과 -1 사이의 하나의 수로 나타냄
- 보통 r로 표시하는 이 계수가 양 극단에 가까우면 상관관계가 강한 것
- IQ와 뇌 부피 사이의 상관관계는 대략 r=0.33 정도로 약한 편
- 뇌의 구분
방법 1. 대뇌(cerebrum)1, 뇌간(brainstem), 소뇌(cerebellum)2으로 나눌 수 있음 (더 세분화해서 9분할 가능)
- 대뇌: 대뇌피질(cerebral cortex), 기저핵(basal ganglia)
- 뇌간: 시상(thalamus), 시상하부(hypothalamus), 중뇌개(tectum), 중뇌피개(tegmentum), 뇌교(pons), 연수(medulla)
- 소뇌
방법 2. 전두엽(frontal lobe), 두정엽(parietal lobe), 후두엽(occipital lobe), 측두엽(temporal lobe)의 네 엽으로 나뉘고, 대뇌의 좌반구와 우반구를 가르는 대뇌종렬(longitudinal fissure)과 전두엽과 측두엽을 나누는 실비우스열(Sylvian fissure), 전두엽과 두정엽을 나누는 중심구(central sulcus)가 존재

- 골상학(phrenology) 피질을 영역들로 구분한 후, 각각의 영역들과 정신 기능은 고정된 연결을 갖고 있다고 하는 학문
- 브로카 영역(Broca’s area): 대뇌 좌반구에 위치한 언어를 구사하는 능력에 특화된 뇌 영역
- 베르니케 영역(Wernicke’s area): 대뇌 좌반구에 위치한 언어 이해 능력에 특화된 뇌 영역
- 위 2가지 영역들은 대뇌 편측화(cerebral lateralization)3의 근거도 제시
2장 경계 논쟁
- 호문쿨루스(homunculus) 운동피질(브로드만 영역 4)과 감각피질(브로드만 영역 3)의 지도
- 중심구의 서로 반대편을 따라 평행을 이룸 (귀에서 귀로 뇌를 관통하는 수직 평면)
- 환상통은 구역 3의 기능이 재배치되면서 생겨남
호문쿨루스(Homunculus)
1. 호문쿨루스(Homunculus) 신경이 밀집된 신체 부위를 비율적으로 나타낸 지도 - 우리 몸의 신경 분포도가 어떠한 식으로 형성되어있는지를 알 수 있음 - 운동신경과 감각신경에 따라 운동 호문쿨
alxalib.tistory.com
- 기능적 MRI (fMRI, functional MRI) 모든 영역의 활동과 뇌의 특정 부위가 얼마나 사용되고 있는지를 보여줌
- 혈류 산소 수준(BOLD, blood oxygen level dependent)4의 신호를 측정함
- 뇌의 특정 부위가 많은 에너지를 사용하며 헤모글로빈을 탈산소화시키면, 사용에 대한 반응으로 해당 부위로 혈액 흐름이 증가
- 각 이미지는 두 개의 유사한 정신 작업에 대응하는 두 이미지를 사용해, 하나에서 배경이 되는 것을 빼줌으로써 얻어짐 (밝게 켜진 영역은 다른 작업에서보다 많이 사용된 영역이라는 뜻, 꺼진 영역도 계속 일하고 있음)

- 뉴런의 수행 능력은 개수보다는 뉴런들이 어떻게 조직되어 있는지에 더 의존함
- 근육의 경우 모두 동일한 방향으로 당기는 작용을 하면 되기에 근육 섬유들이 힘에 미치는 영향은 부가적(additive)임 (실제로, 상관계수는 0.7~0.9)
- 뉴런의 경우 서로 다른, 아주 작은 업무를 수행하며 정교하게 협력함
- 피질 동등 잠재성(equipotentiality): 모든 피질 구역은 서로 다른 기능에 특화되어 있지만, 또한 다른 기능을 담당할 잠재력을 갖고 있음
- 어느 정도는 맞는 말이지만, 피질은 무한히 적응할 수 없음 (적응의 한계)
2부 연결주의
3장 뉴런은 섬이 아니다
- 가중투표 모델 (weighted voting model)
- 수상돌기의 전기신호는 많은 뉴런에서의 신호를 일단 다 받아들임
- 시냅스의 세기는 뉴런이 스파이크를 일으킬지를 결정할 때 그 시냅스가 지니는 가중치(weight)를 나타냄
- 한 뉴런이 다른 뉴런으로부터 여러 시냅스를 받는 중복 투표도 가능
- 투표의 동시성도 중요 (두 시냅스의 투표는 그들의 전류가 시간적으로 중첩될 정도로 거의 동시에 일어날 때만 동일한 선거로 계산)
- 흥분성(excitatory) 시냅스와 억제성(inhibitory) 시냅스가 따로 존재함 (전기 시냅스는 억제성 X)
- 흥분성 뉴런은 다른 뉴런에게 흥분성 시냅스만 만들고, 억제성 뉴런은 다른 뉴런에게 억제성 시냅스만 만듦
- 하지만, 수상돌기에서 받아들이는 시냅스는 흥분성과 억제성 둘 다 가지고 있음
- 따라서, 흥분성 시냅스는 ‘에’(흥분성 신호)라는 신호를 받으면 ‘예’라는 신호를 보내고, 억제성 시냅스는 ‘예’라는 신호를 받으면 ‘아니오’(억제성 신호)라는 신호를 보냄
- 대부분의 뉴런은 흥분성 뉴런임
- 뉴런이 아무 때나 스파이크를 일으키는 것을 방지하는 두 가지 메커니즘: 역치와 시냅스의 억제
- 세포체에서 신호를 다 집계를 하고, 역치를 넘으면 축삭으로 전달됨
- 축삭에서 스파이크 발생여부는 ‘전부 아니면 무’라는 실무율의 법칙을 따름 (역치를 넘어야 함)
4장 밑바닥까지 모두 뉴런
- 내측 측두엽(MTL, medial temporal lobe)에 존재하는 제니퍼 애니스톤 뉴런에 대한 연결주의 해석
- 제니퍼 애니스톤의 얼굴은 단순한 부분요소들의 결합으로 이루어짐 (e.g. 푸른 눈, 금발)
- 뇌에는 이 목록에 있는 각각의 자극을 감지하는 뉴런들을 포함하고 있음 (e.g. 푸른 눈 뉴런, 금발 뉴런)
- 핵심
- 재니퍼 애니스톤 뉴런은 이 모든 부분 뉴런들로부터 흥분성 시냅스를 받음
- 재니퍼 애니스톤 뉴런의 역친느 매우 높아서 모든 부분 뉴런들이 스파이크를 일으켜야만 재니퍼 애니스톤에 대한 만장일치가 이루어져 스파이크를 일으킬 수 있음
- 푸른 눈 뉴런과 같은 부분 뉴런은 까만 동공, 푸른 홍채와 같이 더 세부적인 부분 뉴런들에 의해 자극이 됨
- 위계구조가 있으며, 위계구조를 따라 올라갈수록 뉴런들은 점점 더 복잡한 자극들을 감지함
- 네트워크 배선 규칙
1. 전체를 감지하는 뉴런은 부분들을 감지하는 뉴런의 흥분성 시냅스를 받아들인다 (부분-전체 규칙)
- 다층 퍼셉트론 모델 규칙이기도 함
- 다층 퍼셉트론 모델과 다른 점은 ‘전체에서 부분으로 내려가는’ 하향 신호도 존재한다는 것 (상호작용적 활성화)
2. 부분을 감지하는 뉴런은 그 전체를 감지하는 뉴런에게 흥분성 시냅스를 보낸다
3. 뉴런의 기능은 주로 다른 뉴런과의 연결에 의해 규정된다 (연결주의)
- 기억의 회상
1. 세포군(cell assembly) 흥분성 뉴런들이 시냅스로 상호 연결된 구조
e.g. 목련, 빨간 벽돌집, 첫사랑이라는 관념들이 첫 키스를 회상하는 동안 스파이크를 일으킴 (눈 앞에 목련이 보이게 되면 세포군 내의 다른 뉴런들을 자극하며 결국 첫 키스를 회상하게 됨)
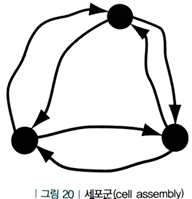
2. 중첩된 세포군
e.g. 목련과 관련되어서 부모님과의 다른 추억이 있음 (목련에 의해서 부모님 추억에 관련된 세포군이 회상될 수도 있음)
- 하지만, 목련 향기가 두 기억을 뒤죽박죽으로 활성화하여 기억을 혼란스럽게 결합할 수 있기 때문에 뇌는 이를 방지하기 위해 각각의 뉴런에게 높은 활성화 역치를 할당할 수 있음
- 뇌의 기억체계는 활성화가 지나치게 퍼져나가면 기억에 혼돈이 생기고, 활성화가 너무 적게 확산되면 전혀 기억이 나지 않기 때문에 여기에 균형을 맞추고 있음 (기억이 결코 완전할 수 없는 이유)
- 억제성 뉴런들은 활성화의 확산을 늦춤으로써 기억 능력을 증가시킬 수 있음
- 중첩된 세포군으로 이루어진 기억인 경우는 회상이 양방향으로 이루어짐
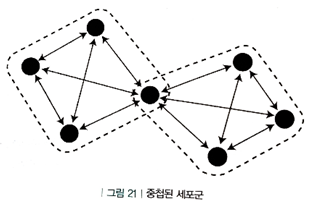
3. 시냅스 사슬(synaptic chain)
- 회상이 한 방향으로만 일어나는, 특정한 시간적 순서를 따라 일련의 사건들이 전개되는 기억
- 이야기 같은 특징을 갖는 기억
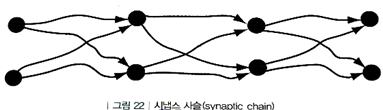
5장 기억의 조립
- 재가중: 시냅스의 세기는 강화될 수도 있고 약화될 수도 있음
- 시냅스는 크기가 커짐으로써 강화됨 (시냅스전은 소포 더 많이 생성 & 시냅스후는 수용체 더 많이 생성)
- 재연결: 시냅스는 생성될 수도 있고 없어질 수도 있음
- 애릭 캔델(Eric Kandel)의 기억과 재가중 연구 ('생명 설계도, 게놈' 16장에 관련 내용 있음)
- 연체동물인 캘리포니아 군소(Aplysia California)는 자극을 받으면 아가미와 흡관을 움츠림
- 자극이 반복되면 그 자극에 더 혹은 덜 민감하게 됨
- 이는 감각기관에서 근육으로 이어지는 신경 경로의 시냅스 세기 변화에 의한 단순한 기억
- 기억이 단 하나의 시냅스에 국한되지 않는, 인간처럼 복잡한 뇌에서는 적용되기 어려움
생명 설계도, 게놈
이후 서술되는 내용은 필자가 매트 리들리의 '생명 설계도, 게놈'을 읽고 인상 깊었던 부분을 정리한 것이다. 1번 염색체 - 생명 - 모든 생물의 98%는 수소, 탄소, 산소로 이루어져 있음 - 최초의 생
alxalib.tistory.com
- 시냅스의 수(재연결)도 학습과 관련이 있기는 하지만, 그 관계가 인과관계인지 아닌지는 분명하지 않음
- 이 책의 필자는 재가중과 재연결로 기억과 관련된 신경연결 패턴들(세포군과 시냅스 사슬)이 생성된다고 제안
1. 두 개의 뉴런이 동시에 반복적으로 활성화된다면, 이들 간의 연결은 양방향으로 강화된다 (동시적 연상)
2. 두 개의 뉴런이 순차적으로 반복하여 활성화된다면, 첫 번째 뉴런에서 두 번째 뉴런의 순서로 연결이 강화된다 (순차적 연상)
- 동시적 법칙과 순차적 법칙은 모두 시냅스 가소성에 대한 헤비안 규칙(Hebbian rules)으로 알려져 있음
- 가소성은 시냅스의 영향을 받는 뉴런들의 활동으로 유발되므로, 이 두 규칙은 모두 활동 의존적
- 헤비안 가소성은 흥분성 뉴런들 사이의 시냅스들만 해당됨
- 망각
- B가 활성화된 동안에 A가 반복적으로 비활성화된다면 A에서 B로의 시냅스가 약화됨
- 시냅스 간의 직접 경쟁으로 연결이 약화되기도 함 (시냅스 생존에 필요한 물질 경쟁)
- 뉴런 하나에는 수만 개의 시냅스가 연결되어 있음 (대부분은 약한 연결)
- 이는 뇌에 1000억 개의 뉴런이 있는 것에 비하면 매우 적은 수 (부피 문제 때문)
- 따라서, 두 뉴런이 연결되어 있지 않은 경우에 연상관계를 학습하는 데 문제가 될 수 있음
- 헤비안 강화로 그 뉴런들을 세포군으로 연결하는 데 성공하지 못할 것
- 그런데, 우리는 한 관념에 대해 하나의 뉴런이 대응하는 것이 아닌 여러 뉴런의 활동에 의해 대응됨
- 따라서, 많은 수의 뉴런들을 사용하다 보면 두 뉴런들의 일부가 우연히 세포군으로 연결될 수 있음
- 신경다윈주의(neural Darwinism)
- 시냅스는 무작위적으로 생성되고, 불필요한 것들은 제거됨 (결국, 시냅스 전체 숫자 유지)
- 기억과 관련된 것들은 살아남고, 더욱 강화됨
- 시냅스 생성은 뇌에게 학습을 위한 잠재력만을 부여하는 그저 무작위적인 과정 (시냅스의 생성 자체는 학습 X)
e.g. 신경근접합은 발달 초기에 하나의 근육에 있는 각 섬유는 여러 축삭에서 오는 시냅스를 받아들이지만, 시간이 지나며 불필요한 시냅스들은 제거되어 각 섬유는 하나의 축삭에서만 시냅스를 받아들임 (더 특정적으로 만듦)
- 이중흔적(dual-trace) 이론
1. 지속적 스파이크 발생을 통한 단기기억 (재가중)
- 재니퍼 애니스톤 뉴런이 하나의 뉴런이 아니라 세포군으로 조직된 한 그룹의 뉴런으로 나타난다고 가정
- 제니퍼 애니스톤이라는 자극이 들어오면, 이 뉴런들은 시냅스를 통해 서로를 계속 자극함
- 세포군의 스파이크는 자기 지속하며 자극이 없어진 후에도 한동안 지속될 수 있음
2. 장기기억으로 전환하기 위해서 뇌는 그 정보를 활동에서 연결로 이전함 (재연결, 세포군이나 시냅스 사슬로)
3. 그 정보를 회상하기 위해서는 그 정보를 다시 연결에서 활동으로 이전해야 함 (그 뉴런들 다시 활성화, 재가중)
- 안정적인 것은 그만큼 가소성이 떨어짐, 그 반대도 마찬가지 (안정성-가소성 딜레마)
- 따라서, 우리는 2개의 기억 저장소를 가지는 것
* 컴퓨터도 RAM(임의 접근 기억장치)과 하드 드라이브라는 두 저장 시스템을 사용
- 문서는 하드 드라이브에 오랜 기간 동안 저장됨 (안정성)
- 문서를 하나 열면, 컴퓨터는 그 정보를 하드 드라이브에서 RAM으로 전송함
- 문서를 편집하는 동안에 RAM에 있는 정보가 변경됨 (램상의 정보는 하드 드라이브에서보다 더 빠르게 수정 가능)
- 문서를 저장하면 컴퓨터는 그 정보를 다시 RAM에서 하드 드라이브로 전송해 보관함
3부 본성과 양육
6장 유전자의 숲 관리
- 뇌의 성장과 발달 4단계 (앞의 두 단계는 태아 시기에 일어남)
1. 뉴런들의 생성
- 이 단계에서 문제가 생기면 소두증(microephaly)을 가지고 태어남
- 마이크로세팔린(microcephalin)이나 ASPM과 같은 여러 유전자의 결함이 원인
- 뇌 크기와 피질은 정상보다 작지만, 접혀 있는 패턴이나 구조상의 다른 특징은 정상에 가까움
- 정신지체를 수반함
2. 뇌 속의 적절한 위치로 이동
- 이 단계에서 문제가 생기면 활택뇌증(lissencephaly)을 가지게 됨
- 임신 기간 동안에 뉴런의 이동을 조절하는 유전자의 변이로 야기됨
- 피질에 주름이 없으며 다른 구조에서도 비정상적인 특징이 나타남
- 심각한 정신지체와 간질을 동반함
3. 가지를 기름 (성장 중인 축삭의 작은 끝부분은 ‘성장 원뿔’로 표현함)
- 뉴런의 표면과 뉴런들 사이 빈 공간은 특수한 유도분자들을 포함함
- 성장 원뿔은 분자 센서를 장착하고 있어 유도분자들에 유도되어 도착지점을 찾음
- 유도분자들과 센서의 생산은 유전적으로 통제됨
- 성장 원뿔이 목적지에 도착하면, 축삭의 가지가 분기함
- 최종 가지의 분기는 유전자의 제어를 받지 않고, 무작위적일 것으로 여겨짐
- 실제로, 뉴런의 전반적 형태는 상당 부분 유전적으로 결정되지만, 상세한 분기 패턴은 거의 무작위적
- 축삭이 적절하게 성장하지 못하면 배선 오류(miswiring)가 발생함 (e.g. 뇌량(corpus callosum) 결손)
4. 연결을 형성
- 유전자는 뉴런의 형태와 그 가지들이 퍼져나갈 영역을 대략적으로 결정함
- 두 뉴런이 포괄하는 영역이 중첩될 경우, 둘 사이에 연결이 이루어질 가능성이 있음
- 하지만, 그들이 실제로 연결될지 여부는 유전자에 의해 결정되지 않음
7장 잠재력 쇄신하기
- 서로 인접한 뉴런들은 유사한 기능을 갖는 경향이 있음
- 또한, 하나의 특정 영역은 제한된 그룹의 입력 영역과 목표 영역에만 배선되어 있음 (다른 영역과 연결 제한)
- 어떤 영역의 기능은 다른 영역과의 연결에 크게 의존
- 댜른 피질 구역과 연결되는 것을 연결지문(connectional fingerprint)라 일컬음
- 연결지문을 이용해 피질 구역이 정의되기도 함 (뉴런의 기능은 주로 다른 뉴런과의 연결로 결정된다!)
- 재배선으로 이 한계를 어느정도 극복할 수 있지만, 민감기 이후에는 유연성이 많이 떨어짐
- 항우울증 약인 플루옥세틴(fluoxetine)은 성인 V1의 가소성을 회복시키는 치료법으로 대두됨
- 해마(hippocampus)와 후각신경구(olfactory bulb)에서는 평생 뉴런들이 계속 추가됨
- 기억이 이루어질 때 내측측두엽에 존재하는 해마에서 먼저 정보를 저장한 후에 이를 신피질과 같은 다른 영역으로 전송함
- 따라서, 해마는 뛰어난 가소성을 가지고 있어야 하며 새로운 뉴런들은 해마에게 추가적인 가소성을 부여함
- 후각신경구는 냄새에 대한 기억의 저장을 돕기 위해 새로운 뉴런들을 사용할 것으로 예상
- 신피질에서는 이미 학습한 것을 유지하기 위해 더 높은 안전성을 필요로 하여 재생성이 일어나지 않음
- 추가로, 신피질의 손상은 새롭게 생성된 뉴런을 끌어당기는 것으로 알려져 있음 (손상된 구역으로 이동)
4부 커넥토믹스
8장 보는 것이 믿는 것이다
- 뇌 3차원 이미지 만들기
- 초마이크로톰(ultramicrotome)5이라는 기계를 이용해 뇌 조직을 얇게 슬라이스냄
- 수조에서 잘라내어 슬라이스가 칼에 들어붙는 문제를 해결함
- 하지만, 물에서 건져낼 때 슬라이스가 접혀버리거나 손상될 문제가 있음 (3번으로)
- 하지만, 그 전에 뇌 조직을 에폭시 수지 안에 넣어 플라스틱 블록으로 만들어야 함
- 뇌 조직은 부드럽고 밀도가 낮아 그냥 자르면 조직이 찢어짐
1. 연속 전자현미경 기술(serial electron microscopy)
- 여러 슬라이스들의 2차원 이미지를 계속 쌓아서 3차원 이미지를 만듦
- 하지만, 이 방법은 슬라이스들이 너무 쉽게 부시지는 문제가 있었음 (2번으로)
2. 연속 블록면 주사 전자현미경 기술(SBFSEM, serial block face scanning electron microscopy)
- 전자가 뇌 조직의 블록에 부딪쳐 튀어나오게 함으로써, 그 표면의 2차원 이미지를 얻을 수 있음
- 그 다음에 초마이크로톰의 칼날로 뇌 조직 블록에서 얇은 슬라이스를 잘라내어 새로운 표면을 드러내고, 다시 그 표면의 이미지를 만듦
- 블록은 단단하기 때문에 쉽게 부서지지 않고, 형태가 쉽게 바뀌지 않아 이미지의 왜곡이 적다는 장점이 있음
3. 자동화된 테이프 수집 초마이크로톰(ATUM, the automated tape-collecting ultramicrotome)
- 컨베이어 벨트처럼 플라스틱 테이프가 물에서 슬라이스를 붙인 채로 밖으로 올라와 건조시킴
9장 자취를 따라서
- 시드니 브레너(Sydney Brenner)의 예쁜꼬마선충 커넥톰 프로젝트
- 당시에는 자르는 기술이 아직 자동화되지 못해서, 몸의 모든 영역이 최소한 한 번은 포함될 때까지 여러 마리의 선충들을 슬라이스로 잘랐음 (현재 밝혀진 예쁜꼬마선충의 커넥톰은 여러 마리가 합쳐진 것)
- 위 과정을 통해 신경계 전체의 모든 시냅스는 알아냈지만, 그 의미를 파악하려면(커넥톰을 찾으려면) 어떤 시냅스가 어떤 뉴런에 속하는지를 구분해야 함
- 당시에는 컴퓨터 기술이 많이 발달하지 않아 수작업으로 연속된 이미지를 분석하여 커넥톰을 완성했지만, 요즘에는 기계학습이 많이 이용됨
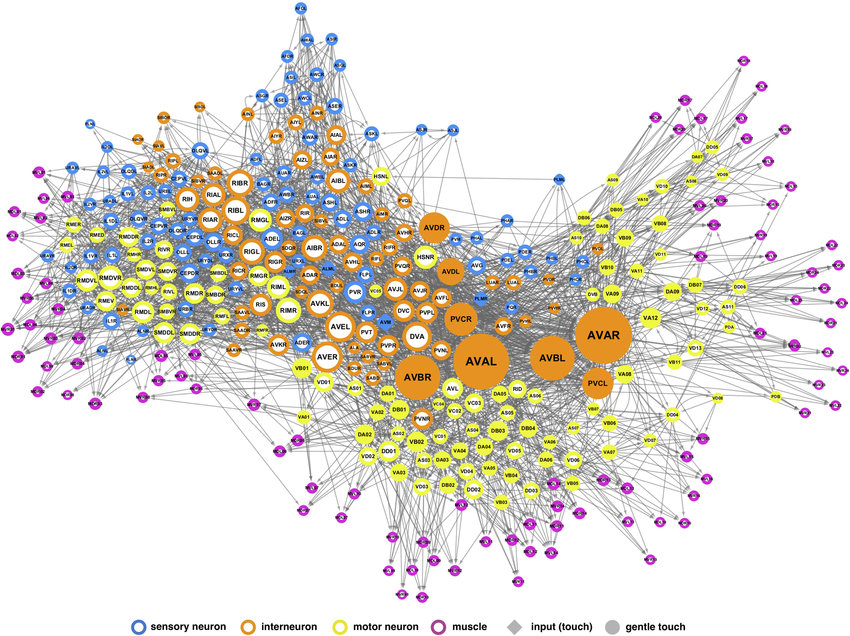
10장 잘라 나누기
- 피질 (층 구조로 이루어짐)
- 신피질(neocortex)/등피질(isocortex)
- 대뇌피질의 대부분을 차지함 (90%)
- 층이 여섯 개로 나뉨 (하지만, 층 구조가 동일한 것은 아님)
- 피질을 잘라낸 위치에 따라 그 층 구조(layering)도 달라짐
- 같은 구역 안에서는 층의 구조가 동일하며 구역 간의 경계에서만 구조가 변함
- 부등피질(allocortex)
- 대뇌피질에서 신피질을 제외 나머지 부분 (10%)
- 여섯 층보다 적은 (혹은 많은) 층을 가지고 있음
- 대표적으로, 해마가 있음
- 뉴런 유형 커넥톰 (neuron type connectome)
- 하나의 유형에 속하는 모든 뉴런을 한 개의 노드로 축약하고, 노드 사이의 선으로 뉴런 유형 간 연결을 표현
- 연결지문을 통해 뉴런의 영역을 구분할 수 있음 (연결은 기능과 직접적인 관계가 있음)
- 예쁜꼬마선충의 302개의 뉴런은 100개 이상의 유형으로 나뉨
- 영역 커넥톰(regional connectome)
- 뉴런의 영역을 구분한 커넥톰
- 기능은 여러 영역 간의 협력으로 이루어지며, 그 협력은 영역 간의 연결로 매개됨
- 브로카 영역(말하는 기능 담당)과 베르니케 영역(언어정보 해석 담당)을 연결하는 궁상다발(arcuate fasciculus)이 존재
- 이 다발들이 손상되면 전달성 실어증(conduction aphasia)6이 발생
- 영역과 뉴런 유형의 커넥톰은 정상적 개인들 간에 큰 차이가 없을 것이며 유전자에 의해 상당 부분 결정될 것
- 이와 대조적으로, 뉴런 커넥톰은 개인별로 큰 차이가 있을 것 (인간의 개별성 이해를 위해 연구되어야 함)
11장 부호 해독하기
- 내측측두엽
- 지각과 생각 모두에 관련되어 있으며, 새로운 서술적 기억(declarative memory)7을 저장하는 데 핵심적인 영역
- 비서술적 기억(nondeclarative memory)에는 운동기술이나 습관 등이 해당하며 선조체(corpus striatum)가 담당함
- 서술적/비서술적 기억 모두 순차적 기억이라면 시냅스 사슬로 뇌에 간직되어 있음
- 해마, 재니퍼 애니스톤 뉴런 등이 여기에 속함
- 해마의 CA3 영역은 서로 간에 시냅스를 형성하고 있는 뉴런들을 포함함 (세포군)
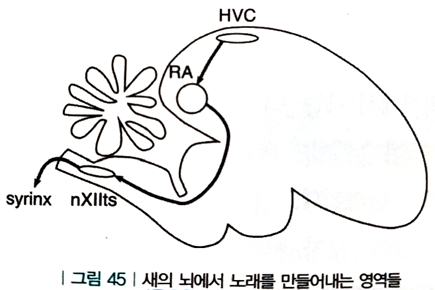
- 새들의 노래 (비서술적 기억)
- 새들이 노래하는 것은 본능적 행동이 아님 (어린 수컷은 몇 달간 아빠 새의 노래를 들으며 연습함)
- 새들은 매번 기본적으로 같은 노래를 부름 (결정화되어 있음, 노래를 기억에 저장하고 회상하는 식)
- 새들은 울대(syrinx)라는 음성기관에 공기를 불어넣어 소리를 냄
- 나오는 소리의 속성들은 울대 주위의 근육에 의해 제어되는데, 그 근육은 뇌에서 오는 지시를 받음
- 새의 뇌에서 전기신호는 HVC에서 RA와 nXII를 거쳐 이동하여 최종적으로 울대에서 소리로 전환됨
- 노래에는 몇 개의 주제부(motif, 0.5~1초 정도 지속)가 존재하고, 각 뉴런들은 자기 주제부가 진행될 때 스파이크를 일으키고 다시 조용해짐 (순차적 스파이크이며 시냅스 사슬에서 유도됨)
- 원형 구조의 경로
1. 새는 스스로 노래를 듣기 때문에, 울대에서 뇌로 되돌아가는 추가적인 단계 필요 (전체 경로 차원)
2. 노래 주제부가 다시 반복되려면 사슬의 마지막 뉴런들은 다시 첫 번째 뉴런들과 연결 (시냅스 사슬 차원)
- 위와 같은 비서술적 기억은 처음에는 어떻게 처음 저장되는 걸까?
- 가능성 1. 재가중
- 어린 수컷의 HVC 뉴런들은 처음에는 무작위적으로 주어지는 입력에 의해 활동함
- 그 중 일부가 헤비안 연결 강화로 강화되고, 이렇게 선택된 서열은 더 자주 활성화됨
- 어떤 하나의 서열이 너무 많이 강화되면 다른 모든 서열을 압도함 (시냅스 생성과 제거는 X)
- 가능성 2. 재가중 + 재연결
- 노래와 연결되지 않는 시냅스 사슬은 약해져 마침내 제거될 것
- 가능성 3. 재가중 + 재연결 + 재배선 + 재생성 (4가지 R 모두)
- 새들이 노래를 부르지 않는 계절에는 HVC가 줄어들지만, 다시 노래를 부르는 계절이 돌아오면 HVC 팽창하는 것 확인함
12장 비교하기
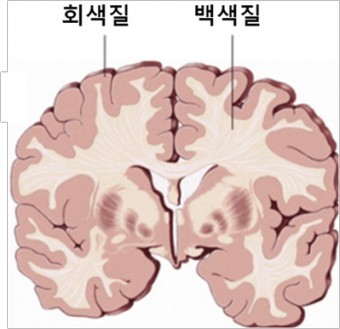
- 회색질(gray matter): 뉴런의 모든 부분들(세포체, 수상돌기, 축삭, 시냅스)로 이루어짐
- 백색질(white matter): 축삭으로만 이루어짐
- 백색질의 축삭들은 대부분 대뇌피질 뉴런들 것이고, 피라미드 뉴런(모든 피질 뉴런의 80%)에 속하는 축삭들임
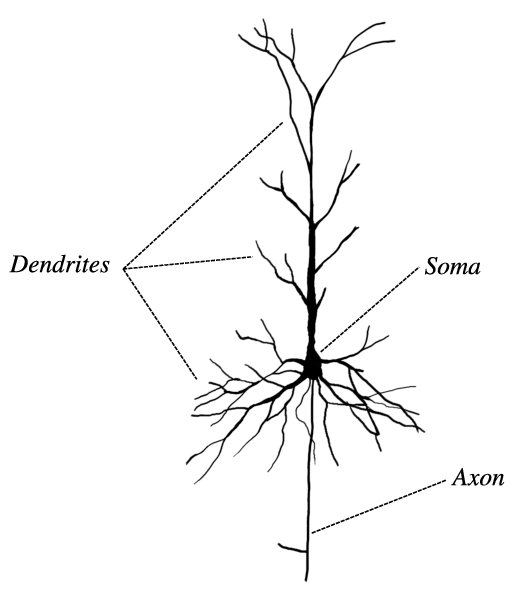
- 피라미드 뉴런(pyramidal cell)
- 피라미드 모양의 세포체와 그 세포체에서 아주 먼 거리를 이동하는 축삭을 가지고 있음
- 피라미드의 꼭대기 부분은 뇌의 바깥쪽을 향해 있으며 축삭은 피라미드의 바닥에서 뻗어져 나옴
- 축삭은 뻗어나가며 옆으로 곁가지(collateral)를 치고 근처의 뉴런들과 시냅스를 형성함
- 축삭의 주가지(main branches)는 최종적으로 회색질을 떠나서 백색질로 들어가 다른 영역의 회색질로 감 (어떤 축삭은 출발했던 근처의 회색질로 다시 들어가기도 함)
- 회색질의 축삭과 수상돌기가 지역 내의 도로라면 백색질의 축삭은 뇌의 고속도로
- 영역 커넥톰을 찾으려면 백색질 내 모든 축삭의 경로를 추적해야 함
- 미엘린(myelin)
- 대부분 지방분자(단백질 30%, 지방 70%)로 구성된 분자이며 척추동물에게만 존재함 (B12가 없으면 만들어지지 않음)
- 수초화(myelination)는 스파이크의 전파 속도를 높여줌 (수초의 지방이 축삭에서 전류의 누출을 방지하는 절연체 역할을 하기 때문)
- 수초화된 백색질의 축삭은 상당히 두껍기 때문에 (두께 1µm 정도) 백색질의 축삭을 추적하는 것만으로도 영역 커넥톰을 찾을 수 있음 (축삭이 가지를 치면 시냅스는 당연히 만들기 때문에)
- 확산 MRI (dMRI, diffusion MRI)
- 트레이서(tracer)를 뇌 내에 주입해 그 위치의 뉴런에서 축삭을 따라 다른 어떤 뇌 영역들로 이송되는지를 시각화하는 MRI (침투적인 기술)
- 트레이서의 운송이 살아 있는 뉴런들의 활동과정에 의존하므로 뇌가 살아 있는 동안에 주입되어야 함
- 공간 해상도가 좋지 않음 (mm 척도의 해상도)
- 가는 섬유 경로는 추적하기 어려우며 경로의 교차 부분에서 정확히 추적하기 힘듦
- 축삭 다발
- 백색질에만 존재하는 구조
- 축삭이 다발을 형성하는 이유
1. 지름길이며 효율적인 경로
- 축삭들은 백색질을 통해 최대한 효율적인 경로를 선택함
- 효율적인 해결책은 종종 하나밖에 없기 때문에 축삭들이 다발을 이루게 됨
2. 앞선 사람 따라하기(follow the leader) 효과
- 뇌의 발달과정에서 축삭은 종종 길을 개척하면서 다른 축삭들이 그 뒤를 따라 올 수 있도록 화학적 단서를 제공한다고 알려짐
- 뇌량(좌반구와 우반구 사이를 오가며 연결), 궁상다발(dMRI로 발견)이 대표적임
13장 변화시키기
- 플루옥세틴
- 세로토닌의 재흡수 억제제 계열의 항우울제
- 플루옥세틴은 세로토닌 수준에 직접적인 영향을 끼치고, 세로토닌의 증가는 장기적으로 뇌에 변화들을 야기함
- 세로토닌은 해마에서 새로운 시냅스, 가지, 뉴런들의 생성을 증가시킴
- 또한, 피질의 재배선을 자극함 (성인의 눈 우세(ocular dominance) 가소성을 회복시킴)
- 성년기에는 새로운 뉴런이 일반적으로 추가되지 않음 (해마와 후각망울(olfactory bulb) 계속 생성됨)
- 뉴런을 생성하는 주요 장소는 뇌실하영역(subventricular zone)임
- 신경모세포(neuroblast)는 성숙하지 못한 뉴런들이며 보통 이곳에서부터 후각망울이라는 뇌 영역으로 이동함
- 뇌졸중이 발병하면 신경모세포의 생성이 증가하며, 그 세포들의 이동 방향을 후각망울 대신 손상된 뇌 영역으로 변경할 수 있음
5부 인간의 한계를 넘어서
15장 ……으로 저장(혹은 구원)하기
- 마인드 업로딩(mind uploading)
- 우리 뇌에 있는 뉴런들의 네트워크를 돌아다니는 전기신호를 시뮬레이션해 최종적으로 자신의 정신을 온전히 컴퓨터에 업로딩하는 것
- 뇌 시뮬레이션과의 입력, 출력을 담당하는 주변 장치(peripheral)들이 잘 만들어진다면, 업로드된 사람은 실제 세계의 모든 즐거움을 누릴 수 있음
- 동일한 유형의 뉴런들은 일반적으로 비슷한 전기적 행동을 보임 (그들의 이온채널이 동일한 방식으로 분포되어 있기 때문)
- 이에 따르면, 신경적 다양성은 유한하며 각 유형마다 모델을 만들 수 있음 (뇌 시뮬레이션에는 커넥톰과 뉴런 유형 모델 두 가지가 필요함)
- 다구획(multicompartmental) 모델
- 뉴런을 여러 구획으로 나누고, 각 이온채널 유형에 하나의 군을 대응시킴
e.g. 하나의 뉴런이 100개의 구획으로 나누어지고 10가지 유형의 이온통로가 존재한다면, 이 모델은 이온채널들의 상태를 규정하기 위한 1000개의 변수들을 포함함 (뉴런에 있는 전체 이온채널의 수보다 훨씬 적음)
- 한 뉴런의 다른 부분들이 독립적으로 기능하는 뉴런들에게 필수적
e.g. 아마크린 세포(amacrine cell): 망막에 위치한 별모양 뉴런으로 수상돌기들이 시각적 움직임의 여러 방향들을 감지하고, 다른 뉴런들에게 서로 다른 신호를 보냄
- 뇌 시뮬레이션은 변화가 이루어질 수 있어야 함
- 변화가 없다면, 업로드된 자아는 새로운 기억을 저장하거나 새로운 기술을 배울 수 없을 것
- 따라서, 뇌 시뮬레이션에 네 가지 R의 모델들을 포함시키는 것이 중요
- 한 가지 난점은 뉴런들이 시냅스의 범위 밖에서 상호작용할 수 있다는 것
e.g. NT는 한 시냅스에서 빠져 나와, 더 멀리 떨어진 뉴런에 의해 감지될 수 있음 (이런 상호작용은 시냅스 밖에서 이루어지므로 커넥톰에 포함되지 않음)
- 전체 뇌 부피의 85%를 차지하며, 대뇌 표면에 몇 mm 두께로 한 겹 덮여 있는 이 조직을 대뇌피질(cerebral cortex) 또는 간단하게 피질이라고 함. 대뇌의 가장 지배적인 부분은 신피질(neocortex)로 약 200억 개의 뉴런 [본문으로]
- 뉴런의 수는 소뇌가 약 700억 개 정도로 제일 많음 (소뇌의 대부분은 과립세포(granule cell)라 불리는 것들로 이 세포들은 매우 작기 때문에 뇌 부피의 10% 정도를 차지) [본문으로]
- 특정 정신 기능은 왼쪽이나 오른쪽 반구 한 쪽이 담당하고 있다는 것 [본문으로]
- 산소화된 헤모글로빈과 탈산소화된 헤모글로빈의 비율 [본문으로]
- 50nm의 두께로 슬라이스들을 자르는 기계 [본문으로]
- 베르니케 영역이 단어들을 받아들일 수 있지만, 받아들인 단어들을 브로카 영역으로 중계 불가 [본문으로]
- 일화적(episodic) 기억과 의미적(semantic) 기억을 포함 [본문으로]
'Books' 카테고리의 다른 글
| 작은 것들이 만든 거대한 세계 (25) | 2024.03.03 |
|---|---|
| 괴델의 불완전성 정리 (24) | 2024.02.13 |
| 인간은 왜 인간이고 초파리는 왜 초파리인가 (57) | 2023.12.18 |
| 도파민네이션 (44) | 2023.08.17 |
| 기억의 뇌과학 (30) | 2023.07.24 |




댓글